REVIEW article Front. Vet. Sc, 26 August 2020 Sec. Veterinary Neurology and Neurosurgery
Acute thoracolumbar spinal cord injury (SCI) is common in dogs frequently secondary to intervertebral disc herniation. Following severe injury, some dogs never regain sensory function to the pelvic limbs or tail and are designated chronically “deep pain negative.” Despite this, a subset of these dogs develop spontaneous motor recovery over time including some that recover sufficient function in their pelvic limbs to walk independently without assistance or weight support. This type of ambulation is commonly known as “spinal walking” and can take up to a year or more to develop. This review provides a comparative overview of locomotion and explores the physiology of locomotor recovery after severe SCI in dogs. We discuss the mechanisms by which post-injury plasticity and coordination between circuitry contained within the spinal cord, peripheral sensory feedback, and residual or recovered supraspinal connections might combine to underpin spinal walking. The clinical characteristics of spinal walking are outlined including what is known about the role of patient or injury features such as lesion location, timeframe post-injury, body size, and spasticity. The relationship between the emergence of spinal walking and electrodiagnostic and magnetic resonance imaging findings are also discussed. Finally, we review possible ways to predict or facilitate recovery of walking in chronically deep pain negative dogs. Improved understanding of the mechanisms of gait generation and plasticity of the surviving tissue after injury might pave the way for further treatment options and enhanced outcomes in severely injured dogs.
– “Deep pain negative”: term synonymous with “absent pain perception” and defined as an absent behavioral response to noxious stimulation caudal to the injury level. For thoracolumbar SCI, this refers to absent pain perception to a mechanical stimulus in the medial and lateral toes of both pelvic limbs and base of the tail; in dogs, this term is applied in the acute setting with concurrent paraplegia to imply a functionally complete injury though sensory and motor status should be considered separately in the chronic setting.
– Sensorimotor complete injury: term used to describe functionally and/or physically complete injury in people where there is absent voluntary movement or pain perception below the injury level; synonymous with AIS-A designation using human SCI grading parameters.
– Ambulatory: the ability to rise and take at least 10 consecutive weight bearing steps unassisted without falling.
– Chronically paralyzed: broad, non-specific term used to capture the population of dogs with permanent neurologic impairment (motor, sensory, and/or deficits in continence) following severe SCI. Dogs in this group can exhibit paraplegia (i.e., no pelvic limb movement at all) or display varying degrees of pelvic limb movements that fall short of being useful (i.e., they remain non-ambulatory).
– Spinal walking: independent ambulation in a “deep pain negative” dog typically characterized by lack of coordination between thoracic and pelvic limbs, difficulty turning, or going backward, intermittent falling (especially when changing directions), frequently intact toe knuckling response but absent hopping, and increased spasticity.
The majority of dogs suffering from acute spinal cord injury (SCI) will recover adequate or even normal function (1). However, a subset of dogs with severe injury fail to regain pain perception caudal to the injury level (“deep pain negative”), remain incontinent and are classified as having an unsuccessful outcome (1–3). The permanent lack of pain perception has been commonly, and frequently incorrectly, interpreted as an indication of spinal cord transection, complete disconnection from all supraspinal influence and minimal to absent chance of meaningful recovery of function. However, a proportion of permanently deep pain negative dogs demonstrate notable spontaneous motor recovery over time (2, 4, 5). This can range from non-purposeful kicking movements of the limbs, especially following tactile stimulation below the injury level, to overground walking with minimal apparent paresis or ataxia. Ambulation exhibited by this population, typically known as “spinal walking,” is commonly considered exclusively reflexive stepping generated by the spinal cord caudal to the level of injury as described by experimental studies of SCI in dogs and other species (6–14). While relatively autonomous circuits within the spinal cord are integral, gait generation is a complex process with extensive coordination between various components of the central and peripheral nervous systems. Understanding the ways in which this circuitry is altered and also how it can recover after injury has broad therapeutic and translational implications.
This review will provide a comparative overview of locomotion and explore the physiological underpinnings of “spinal walking” after severe SCI in dogs. Additionally, the clinical characteristics of motor recovery with absent pain perception as well as proposed means to predict and facilitate its development in this population will be described.
Normal locomotion is a complex action that involves coordination of multiple brain regions, circuitry within the spinal cord and peripheral nerves and muscles. The basic components of locomotion are evolutionarily conserved with broad overlap even between invertebrate and vertebrate animals. While motor systems within the brain and spinal cord are essential to producing locomotion, integration of sensory input at all levels is also integral to proper functioning and modulation of locomotion in response to environmental surroundings.
Within the brain there are several motor regions from which upper motor neurons arise to produce the descending motor tracts, with some variability in their relative importance across species. These include the primary motor cortex located in the parietal lobe of each cerebral hemisphere, the red nucleus of the midbrain and the reticular formation of the pons and medulla oblongata. Additionally, the mesencephalic locomotor region located just ventral to the caudal colliculi is involved in initiating stepping movements. Axons of neurons from this area do not directly project to the spinal cord but rather interact with other brainstem motor regions, especially within the reticular formation, to produce locomotion. All of these components are also influenced and modulated by the cerebellum and basal nuclei. Input from these areas allow for complex movements and adjustment of locomotor activity. The overall output of the brain activates spinal cord motor circuitry and produces voluntary motor activity (15–18).
Axons of the upper motor neurons (UMN) in the various motor regions of the brain form the descending motor tracts to the spinal cord. These descending motor tracts produce both inhibitory and excitatory influence on spinal cord interneurons and lower motor neurons (LMN) to initiate and regulate voluntary movement. These include the lateral and ventral (the latter being more developed in primates) corticospinal tracts, rubrospinal tract, and pontine and medullary reticulospinal tracts. While the vestibulospinal tracts play a crucial role in posture and influence locomotion, they will not be discussed in detail. The corticospinal tract originates in the primary motor cortex, follows the major descending white matter pathway (internal capsule, crus cerebri, pyramids) to the medulla where the majority of fibers cross at the pyramidal decussation to descend in the lateral funiculus of the contralateral spinal cord. It is primarily involved with complex and precise movements although is reported to retain a role in overall gait generation (17, 19, 20). The rubrospinal tract originates in the red nucleus, immediately crossing midline to travel in the contralateral lateral funiculus of the spinal cord. The pontine and medullary reticulospinal tracts start in the ill-defined reticular formation of the brainstem before descending in the ipsilateral ventral and lateral funiculi, respectively. The rubrospinal and medullary reticulospinal tracts facilitate flexor muscles and inhibit extensors while the pontine reticulospinal tract does the opposite, providing a tonic balance between facilitation and inhibition of spinal cord lower motor neurons (15–18). Direct evidence in dogs is sparse, but it has been demonstrated in cats that the reticulospinal pathways play an important role in postural control and basic gait generation on a flat surface while the rubrospinal tract is involved in both normal control of locomotion and in producing adaptive movements to changes in the environment (19, 21). The corticospinal tract is less well-developed in domestic species (compared to people and non-human primates) and is not considered essential to generate basic locomotor rhythms; however, it functions in parallel with the other motor pathways to primarily regulate and fine tune movements (19–21).
Within the spinal cord, circuitry involved in gait generation has been identified in multiple species and is known as the central pattern generator (CPG) (13, 16, 18, 22–24). The CPG organizes the basic pattern for stepping, independent of supraspinal or sensory input. This basic rhythmic pattern of the CPG is produced by interconnected, alternating, and mutually inhibitory flexor and extensor interneurons (25). This network is thought to extend the length of the spinal cord but has been most extensively studied in the lumbar region in relation to control of the pelvic limbs (in quadrupeds, or legs in people). In this context, it is located in the intermediate zone of spinal cord gray matter although the precise cranial to caudal location of integral components of the circuitry within the lumbar spinal cord might vary between species (e.g., cranial lumbar in dogs, rats, people; mid-caudal lumbar in cats) (24, 26). These interneurons, in turn, activate lower motor neurons via additional intermediary interneurons, the output from which serves as the final common pathway to produce locomotion via direct innervation of appendicular muscles (25). The CPG also provides coordination between left and right limbs via integration of commissural interneurons and thoracic and pelvic limbs, important in normal quadrupedal locomotion (15, 18, 27, 28). While autonomously capable of relatively complex patterns of activity, under normal (non-injured) conditions, supraspinal input is necessary for activation (29). Additionally, modifying input to the CPG is necessary to allow adaptation of the basic alternating stepping pattern. Sensory input derived from visual information, vestibular input, and both exteroceptor and proprioceptors located on the body and limbs is also an important component of locomotion, specifically providing information needed to adapt locomotion to an animals surroundings (16).
Gait generation itself consists of two major phases, the postural stance phase and the protraction or swing phase. However, based on the activation pattern of specific pelvic limb muscles, the step cycle should really be considered as having four phases: flexion and first extension occur during swing while second and third extension occur during stance (30, 31). Second extension happens during the early part of the stance phase when the knee and tarsus joints actually flex despite contracting extensor muscles as the animal prepares to bear weight (30, 31). Third extension is characterized by hip, knee and tarsus extension as the weight of the body is pushed forward (30, 31).
Plasticity of Locomotor Systems After SCI
The central nervous system is largely considered to have poor regenerative capacity; however, remarkable plasticity is possible. In fact, much of what is known about the organization and function of locomotor systems has been elucidated via various experimental spinal cord transection and decerebrate animal models (6, 7, 9–14, 32). Reorganization and adaptations that occur at all levels might influence recovery of motor function below the level of severe injury. These include regrowth of axons across the epicenter, recovery/reactivation of conduction of residually intact UMN axons traversing the lesion epicenter, a more autonomous role for the CPG, alterations in excitability of interneurons and LMNs below injury, activation of silent synapses, changes in synaptic weight, and alterations in sensory input or how afferent input is integrated at the level of the spinal cord below injury (29, 33–38).
Axonal regeneration of UMN axons has been demonstrated via experimental transection models although the capacity for regeneration varies between axon types and is limited compared to axons in the peripheral nervous system (37, 39–41). While serotonergic axons have demonstrated robust sprouting ability after injury, there are substantial deterrents to meaningful regrowth of most other disrupted axon systems (41). These include the size of the defect, astroglial scar formation, growth inhibitory molecules (e.g., chondroitin sulfate proteoglycans) and myelin-based growth inhibition (37, 39, 40). Additionally, there is no guarantee that regenerating axons will reconnect with the appropriate below-injury targets. These factors lead to minimal functional recovery in most complete transection models. There is active research regarding how to facilitate more effective regrowth through the use of various grafts, scaffolds, inhibitors of scar formation and other modulators of axonal growth (37, 39, 40, 42–47).
Fortunately, even with severe injury, physical spinal cord transection is uncommon. Residually intact, small diameter, subpial UMN axons traversing the lesion epicenter have been shown in various animals and people with functionally complete injury (33, 35, 48, 49). While the degree of loss of large diameter axons and abnormal myelination of residual fibers contribute to persistent neurologic deficits in chronic SCI, there is evidence of reactivation of surviving long tract axons within rubrospinal and other descending motor tracts (50, 51). This might serve to reestablish supraspinal influence on spinal cord circuitry and LMNs and contribute to recovery of voluntary motor control (35, 50). Prior work in rats and cats has shown that as little as 5–10% of the original population of axons can allow voluntary ambulation after severe injury (33, 50, 51).
Additionally, collateral sprouting of spared UMNs and regrowth of local propriospinal fibers traversing the site of injury have each been shown in experimental injury in rodents and lampreys (52–55). These mechanisms serve to produce novel, multisynaptic pathways, and reestablish the connections between UMNs and LMNs with associated improvements in motor function (39, 52). Interestingly, propriospinal neurons have also been shown to activate CPGs, highlighting their potential importance in achieving useful locomotor recovery after severe injury (39, 55–57).
Below the level of injury, notable changes also occur. There is increased importance of the integration between sensory input and CPG activity to coordinate motor output due to limited or lack of supraspinal control (29). Alterations in both motor neuron pool excitability and sensory input to the dorsal horn occur and likely contribute to functional status after injury (38, 58–62). For example, pharmacologic inhibition of post-synaptic inhibition with strychnine has been used to facilitate spinal walking in experimentally transected dogs (63). However, maladaptive plasticity and development of aberrant neuronal circuits commonly manifested as neuropathic pain or spasticity can also occur and impair functional recovery (58, 60).
The Spinal Cord
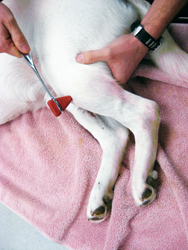
Photo courtesy of Dr. Phil Zeltzman.
The cranial tibial reflex causes flexion of the hock.
Incidentally, these four stages correlate with the anatomy of the spinal cord. Nerve fibers involved with proprioception are located superficially in the cord. This explains why a mild lesion has mild effects on the patient.
A deeper lesion will affect the nerve fibers that control motor function. And a very severe lesion will apply pressure in the deepest nerve fibers—those that relay deep pain.
These four stages help us determine the severity of the lesion. A dog with proprioceptive deficits is mildly affected. At the other end of the spectrum, a dog with no deep pain, to simplify, may have a poor prognosis.
Patients may go through these four stages very slowly—over weeks or months—or within days or even hours.
No pain sensation in the hind paws Pt 2
The neurological exam seems to be one of the most confusing concepts in veterinary medicine. Let’s try to clarify things. To simplify, we will focus on the hind legs only.
One way to look at the neuro exam is to divide it into an assessment of “the four stages” and an evaluation of four reflexes.
The four stages require a functional spinal cord and a functional brain, and they help us determine a lesion’s severity. Reflexes tell us only where the lesion is localized.
Since I am a surgeon and not a neurologist, I talked to Todd Bishop, DVM, Dipl. ACVIM (neurology), of Upstate Veterinary Specialties in Latham, N.Y., to ensure the accuracy of the following information.